Para cualquiera de nosotros es bien conocido que la historia de una nueva vida humana empieza con una fecundación, o sea, con la unión de dos células (gametos): óvulo y espermatozoide. Los gametos son haploides (n), es decir, tienen un único juego de cromosomas (en nuestro caso, 23), de forma que tras la fecundación, el cigoto resultante es diploide (2n) y recupera el doble juego de cromosomas que caracterizan al organismo de turno (en la especie humana, 46). Este proceso tan sencillo es la norma, no exenta de excepciones, en el reino animal. Sin embargo, a menudo nos creemos que lo que es válido en nuestro caso se puede extender como algo general y esto no es así. Por ejemplo, en las plantas con flores y frutos (angiospermas) se da un proceso bastante más rebuscado que ha tenido consecuencias muy relevantes para nuestra vida diaria, y podríamos decir que el mundo que vemos hoy es como es precisamente por esta complicación. En las angiospermas, la historia de una nueva vida comienza con una insólita doble fecundación.
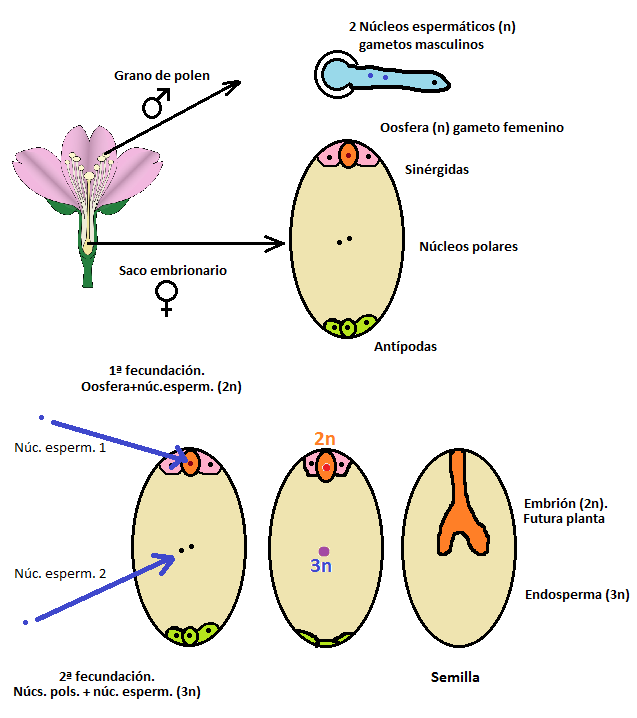
Los que estudiamos biología, en algún que otro momento tuvimos que aprendernos, casi seguro que de memoria y sin comprender muy bien a cuento de qué, el dibujo de abajo. Representa el equivalente a la semilla antes de ser fecundada, lo que suele llamarse saco embrionario: un conjunto de siete células y ocho núcleos haploides que parece diseñado para suspender exámenes de botánica. En uno de los polos está la oosfera (el gameto femenino propiamente dicho), rodeada de dos células sinérgidas (que ayudan en la fecundación). En el centro, acaparando casi todo el citoplasma hay dos núcleos polares, y finalmente en el extremo opuesto hay tres células llamadas, precisamente, antípodas. Llegada la hora de la verdad, el grano de polen aporta no uno, sino dos núcleos espermáticos haploides, que hacen de gametos masculinos. Uno de ellos fecunda la oosfera formando una célula diploide y el subsiguiente embrión, es decir, la futura planta. Hasta aquí lo normal y esperable, vaya. El otro núcleo espermático, sin embargo, se une a los dos núcleos polares. Esta segunda fecundación origina por lo tanto un insólito núcleo triploide (3n), que acaba generando lo que se conoce como endosperma, el “relleno” de la semilla.

¿Qué tiene de especial este endosperma triploide? Los cariotipos triploides son, ya de por sí, algo muy excepcional, normalmente debidos a divisiones celulares fallidas. Sin embargo aquí tenemos una situación muy concreta en la que un tejido es habitualmente triploide y que sólo se da en angiospermas. Se trata de un tejido nutritivo destinado a alimentar al embrión durante la germinación, y al ser característico del grupo de plantas que más éxito han tenido en la Tierra desde que surgieron en el Cretácico, es razonable preguntarse si el endosperma triploide ha supuesto alguna ventaja para su ciclo vital. Podría pensarse, por ejemplo, en que favorece una maduración más rápida de la semilla (y ciertamente, las angiospermas llegan a ser mucho más rápidas en este aspecto que sus precursoras, las gimnospermas). Lo que es indudable es que para nosotros mismos el endosperma triploide resulta esencial en nuestra vida. Un rápido repaso nos permitirá darnos cuenta de que la mayor parte de la ingesta calórica de la humanidad depende, directa o indirectamente, de este endosperma triploide de las angiospermas: a los cereales, que dan de comer al mundo entero (el arroz, el trigo, el maíz y todos los demás) hay que añadir las legumbres y los frutos secos, de los que también obtenemos calorías fundamentalmente a partir del endosperma de las semillas. Pero es que si además consideramos que en gran medida los animales domésticos se alimentan a su vez de semillas o piensos procesados a partir de las mismas, no debería parecernos una exageración admitir que este tejido concreto está dando de comer al mundo entero.
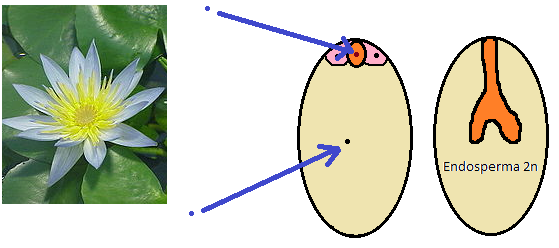
Un par de preguntas razonables que podemos hacernos ahora es cuándo y cómo evolucionó este tejido tan relevante con esta dotación cromosómica tan excepcional. A esta y otras preguntas lleva dedicándose Ned Friedman y su equipo de embriología vegetal de la universidad de Harvard, que hace unas semanas dio una conferencia en el Real Jardín Botánico de Madrid. Uno de los objetos de estudio del equipo de Friedman han sido las familias que, según las hipótesis filogenéticas más recientes, están más cerca de la base de la evolución de las angiospermas, por ejemplo, los nenúfares (Nymphaceae). El saco embrionario de los nenúfares es muy distinto al caso común que hemos visto antes.

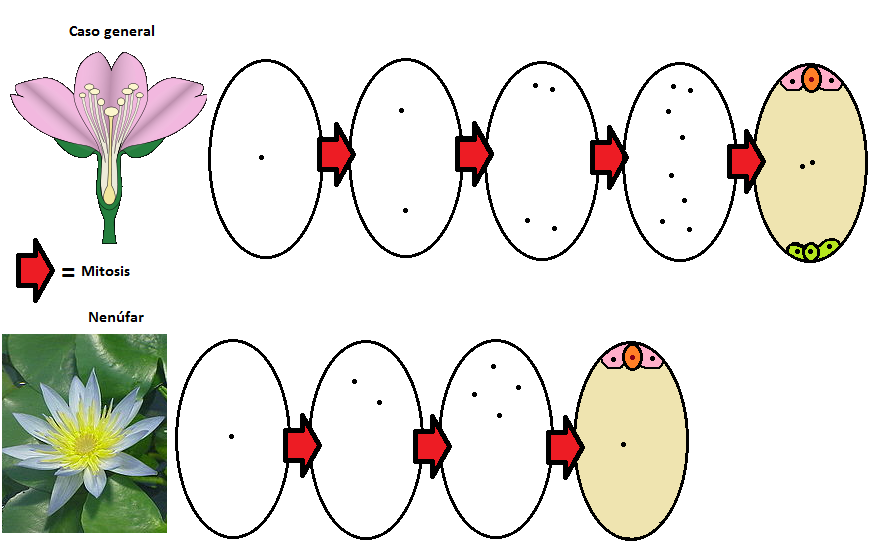
No se aprecian dos núcleos polares, sino sólo uno, y las antípodas están ausentes. Los libros de embriología clásicos interpretaban esta situación alegando que por algún motivo los núcleos polares se fusionaban prematuramente y que las antípodas degeneraban. Sin embargo lo que de verdad estaba ocurriendo era mucho más sencillos: esos núcleos no se observaban porque nunca estuvieron allí. El desarrollo de las semillas de los nenúfares, relacionados con las angiospermas más primitivas, es diferente: en lugar de ocho núcleos y siete células hay cuatro núcleos y cuatro células, y su endosperma es diploide, y no triploide.

Sorprendentemente, que el famoso endosperma triploide no estaba presente en el mismo origen de las plantas con fruto, sino que evolucionó en algún momento determinado entre las angiospermas primitivas. ¿Cómo? Podría pensarse que el aumento de complejidad que se aprecia entre el saco embrionario de un nenúfar y el de la mayoría de las angiospermas no pudo surgir sin una evolución compleja…

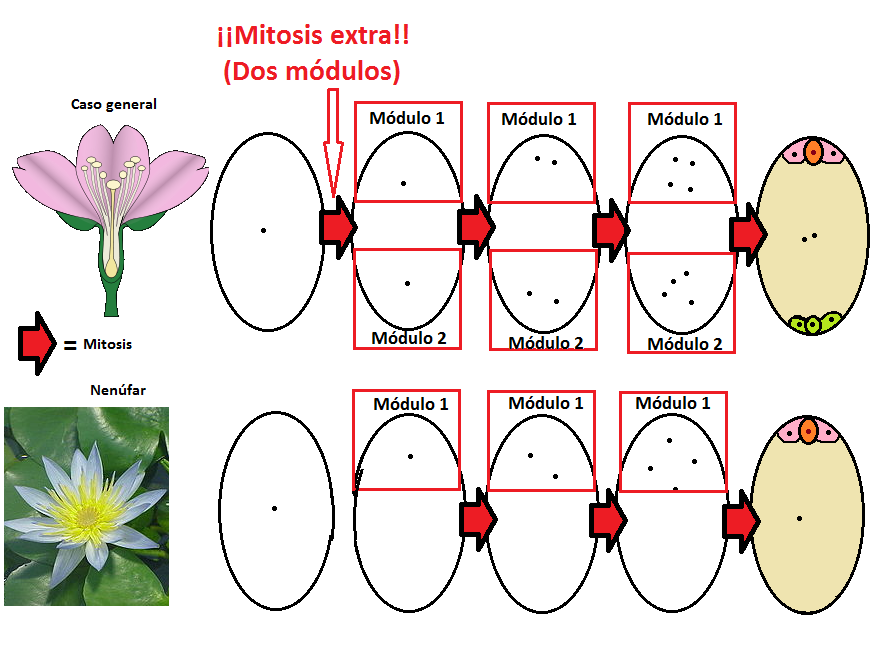
…pero en realidad, si se atiende al desarrollo de ambas configuraciones, se puede llegar a la conclusión de que ambos casos son muy similares y que responden simplemente a una duplicación del módulo básico del caso del nenúfar. Es decir, la evolución del endosperma triploide se explica con la aparición de un módulo adicional de cuatro núcleos, de forma que en el centro del saco quedan dos núcleos polares en lugar de uno, como era la situación primitiva. Si nos fijamos, esto se debe a algo tan sencillo como una mitosis adicional al comienzo del desarrollo de la estructura. Tan simple como eso.

Un dato que corrobora esta hipótesis es que existen otras excepciones interesantes en algunas angiospermas, como en la familia de las peneáceas (Penaeaceae), características del Reino Capense. Algunas de estas plantas tienen sacos con cuatro núcleos polares, y tras la doble fecundación producen endospermas pentaploides (5n). Efectivamente, se trata de sacos embrionarios con cuatro módulos.

Hay muchos factores que han determinado el éxito evolutivo de las angiospermas (entre otros, la coevolución con los insectos), pero muy posiblemente la aparición del endosperma triploide fue uno de ellos. Lo interesante del estudio del desarrollo modular de Friedman reside en que la explicación del cómo y el cuándo surge este rasgo tan curioso es sencilla: una mitosis extra intercalada en el desarrollo del saco embrionario, que sin embargo ha tenido apetitosas consecuencias.
Rafael Medina
Referencias
Friedman, W., Madrid, E., & Williams, J. (2008). Origin of the Fittest and Survival of the Fittest: Relating Female Gametophyte Development to Endosperm Genetics International Journal of Plant Sciences, 169 (1), 79-92 DOI: 10.1086/523354
Friedman, W., & Williams, J. (2003). Modularity of the Angiosperm Female Gametophyte and Its Bearing on the Early Evolution of Endosperm in Flowering Plants Evolution, 57 (2) DOI: 10.1554/0014-3820(2003)057[0216:MOTAFG]2.0.CO;2
Tomado de:
Hablando de Ciencia